O desenvolvimento da planta ocorre de forma orquestrada pelo desenvolvimento celular programado e pela sua modulação por sinais ambientais como temperatura, disponibilidade de água, fotoperíodo e qualidade/intensidade luminosas. Esses sinais ambientais são “lidos” por estruturas especializadas nas plantas – e.g. pigmentos e receptores – e convertidos em sinais químicos, como os hormônios vegetais, no interior das células. Esses hormônios, por sua vez, disparam mensageiros secundários que induzirão respostas fisiológicas no núcleo da célula.
Está sem tempo para ler? Não tem problema, nós preparamos uma versão em áudio deste post para você ouvir.
De forma geral, essas respostas dependem de um estímulo mínimo para que ocorram. Por isso, a concentração e o balanço entre os hormônios, bem como a presença e o número de receptores para aqueles hormônios nas células alvo, são elementos importantes para que uma resposta fisiológica seja desencadeada.
A modulação do crescimento das plantas é feita pelo balanço equilibrado de diferentes hormônios vegetais que agem sinergicamente promovendo ou inibindo o desenvolvimento vegetal, das sementes ao indivíduo adulto. Os hormônios notadamente promotores do crescimento vegetal são as auxinas (AX), citocininas (CK) e giberelinas (GA), enquanto que ácido abscísico (ABA) e etileno (ET) reduzem ou inibem o desenvolvimento por diferentes processos, sendo o fechamento estomático promovido por ABA e a senescência precoce decorrente da produção de ET em níveis supra ótimos os principais limitantes para a manutenção da fotossíntese e do potencial produtivo.
Ressalta-se que mesmo em condições controladas, o etileno é sintetizado durante todo o ciclo de vida das plantas, ao passo que o ácido abscísico é produzido quando há perda de potencial de turgor em células da raiz ou da folha. Um estudo conduzido pela NASA demonstrou que o ET é emitido em altas concentrações durante o período reprodutivo da cultura da soja, paralelamente à queda dos níveis de CK demonstrado por Carlson, levando ao abortamento natural de flores (Figura 1).
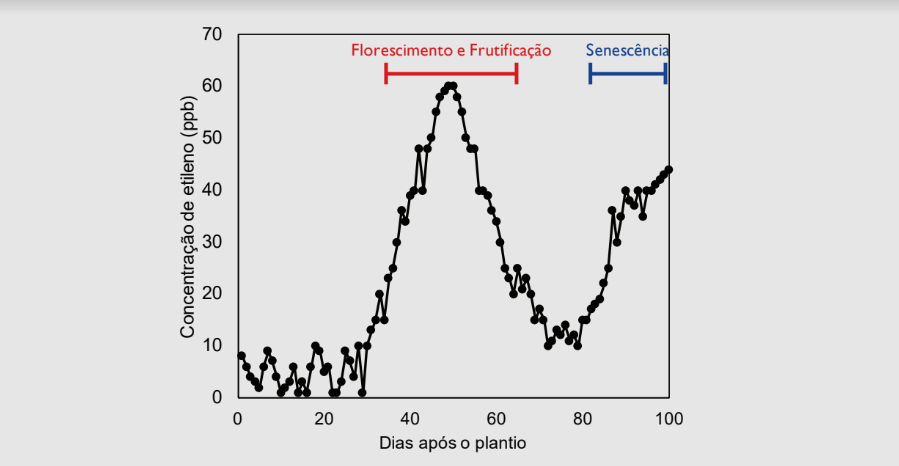
Qual a principal causa do abortamento floral em soja? Em média, apenas 6% das flores da soja não são fertilizadas, ou seja, em apenas uma pequena o grão de pólen não é capaz de traçar o seu caminho até o óvulo (Abernethy, Palmer, Shibles, & Anderson, 1977). Se não há problema de fertilidade, por que isso ocorre? Baixe este e-book grátis e entenda!
Dada a essencialidade dessas substâncias para o crescimento das plantas, foram destacadas abaixo as principais funções dos hormônios vegetais promotores, no que diz respeito à modulação morfológica das plantas.
Hormônios Vegetais
Auxina
Durante o desenvolvimento, as AXs são produzidas especialmente nos meristemas apicais caulinares e transportadas de forma ativa por transportadores de membrana no sentido das raízes. Conjuntamente com as GAs, promovem o alongamento de células jovens da parte aérea e do sistema radicular. Ressalta-se que balanços entre AX e CK voltados para AX promovem maior desenvolvimento de raízes e manutenção das gemas laterais caulinares em estado de dormência, resultando em parte aérea menos desenvolvida (Figura 2).
Citocina
As CKs, por sua vez, são sintetizadas majoritariamente em meristemas apicais radiculares e transportadas via xilema para a parte aérea. Esse hormônio tem papel fundamental no controle das etapas da divisão celular nos tecidos vegetais e na quebra de dormência de gemas axilares. De forma contrária ao balanço visto anteriormente, balanços entre AX e CK voltados para a CK promovem quebra da dormência de gemas laterais e maior desenvolvimento da parte aérea em detrimento ao desenvolvimento de raízes (Figura 2).
Giberelina
A giberelina é sintetizada em todo tecido jovem em desenvolvimento. Esse hormônio é capaz de controlar a organização do “esqueleto” celular, organizando microfibrilas de celulose de forma que a célula possa expandir-se longitudinalmente. Essa expansão em comprimento é que define o comprimento dos entrenós das plantas e é importante na definição da arquitetura do caule (Figura 2).
Vale lembrar que os fitormônios possuem outras funções no metabolismo das plantas além das acima mencionadas. Outras funções serão discorridas em materiais futuros.

A importância do balanço entre os hormônios vegetais para o desenvolvimento da planta.
A utilização de reguladores vegetais – moléculas sintéticas capazes de promover respostas similares aos hormônios vegetais – é prática comum à agricultura Brasileira há mais de 20 anos visando a maximização das produtividades agrícolas. Como pôde ser visto, a utilização de reguladores vegetais isolados (Figura 2) não foi eficiente na promoção do crescimento equilibrado de plantas de soja, visto que:
- A pulverização com auxina promoveu o maior desenvolvimento de raízes, mas com um custo significativo à parte aérea quando comparado ao tratamento Stimulate.
- Plantas tratadas somente com citocinina engalharam mais, mas o menor desenvolvimento do sistema radicular comprometeu a reposição de água durante a transpiração, possivelmente mantendo os estômatos fechados por maior período. A menor condutância estomática reduziu a difusão do dióxido de carbono na câmara subestomática, limitando a fotossíntese e o desenvolvimento da planta como um todo.
- A giberelina promoveu o estiolamento de caule e pecíolos, resultando em uma planta com aspecto de cipó, sem sustentação.
Essas características morfológicas, associadas aos incrementos fotossintéticos favorecidos pela citocinina e giberelina a nível molecular, resultam em maior síntese de carboidratos e de plantas com maior potencial produtivo. Além disso, a utilização de Stimulate e Hold durante o florescimento favorece a produtividade agrícola, pois complementa os níveis endógenos de CK – que se encontram naturalmente baixos durante essa fase além de inibir a síntese de etileno, que conforme mostrado na figura 1, estão elevado durante o florescimento.
Dinâmica dos hormônios vegetais durante a germinação
Nas sementes estão presentes toda a energia e recursos necessários para o crescimento inicial do embrião até a formação das raízes e folhas primárias. Após esse período as plantas recém emergidas tornam-se autotróficas e dependentes da energia e dos recursos existentes no ambiente para dar continuidade ao seu crescimento e desenvolvimento.
A partir da embebição do tegumento, há ativação da maquinaria metabólica do embrião, para que o ácido abscísico – que mantém a semente dormente – seja degradado. O concomitante aumento dos níveis de giberelina promovem a germinação, pela ativação de enzimas responsáveis pela quebra de açúcares complexos em açúcares simples, que nutrirão a plântula. Neste momento ocorre uma ação coordenada e conjunta de outros dois hormônios: a citocinina, responsável pela divisão celular, que juntamente à giberelina promoverá o alongamento do hipocótilo, e a auxina, que junto à giberelina, promoverá a protusão e o desenvolvimento da raiz primária.
Em gramíneas a citocinina também é responsável pelo aumento da permeabilidade da membrana da camada de aleurona.
































0 comentários